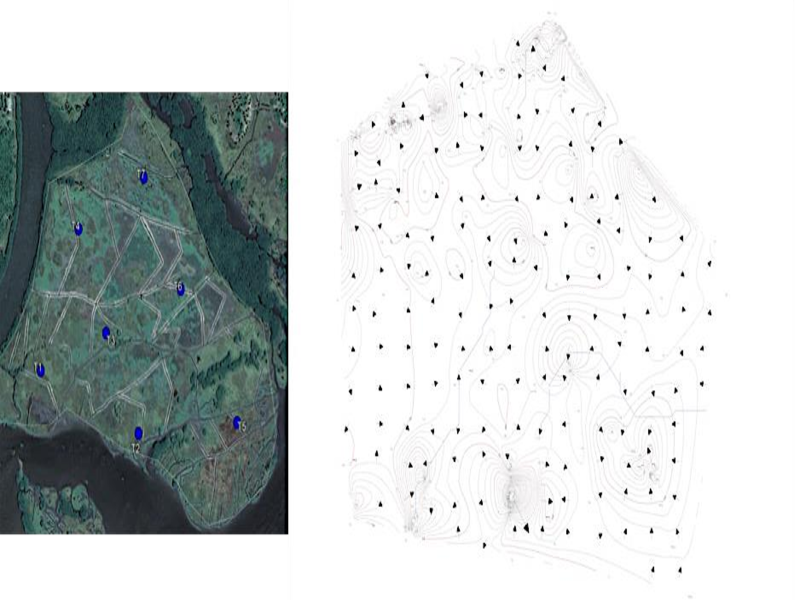
El presente proyecto tiene como finalidad establecer un programa de restauración ecológica integral en una superficie de 13 hectáreas en dos sitios de Costa Rica y 30 ha en Benín, África durante el periodo del 2017 al 2023; transfiriendo la tecnología aplicada en programas de restauración ecológica en México. La restauración se efectúa mediante el manejo hidrológico, a través de la aplicación de modelos hidrodinámicos y el análisis químico del agua intersticial (calidad de agua) que garantice la dinámica hidrológica y la recuperación de la cobertura vegetal, mediante la reforestación de propágulos de Rhizophora mangle, Rhizophora racemosa, Avicennia germinans y Pelliciera rhizophorae. Esto mediante la cantidad mínima de muestreos, para calibrar los modelos numéricos capaces de describir la dinámica del agua y los sedimentos del agua intersticial, para posteriormente evaluar la respuesta de dicha dinámica (corrientes, hidrología, calidad del agua y sedimentos), ante diferentes escenarios de manejo hidrológico, concentración de la salinidad y los nutrientes, así como del potencial redox y la respuesta de la regeneraciónnatural.
El Rector de la Universidad Autónoma de Campeche, en conjunto de la Abogada General, la investigadora Dra. Claudia M. Agraz Hernández y el técnico Ing. Jordán Efrén Reyes Castellanos asistieron a la inauguración del proyecto. Las autoridades que se presentaron al evento fueron la alcaldesa de Ouidah, la señora Celestine Adjanohoun. Las actividades que se llevaron a cabo en alcance al evento fue la misma inauguración, visitas a las comunidades donde se produce la sal, bosques de mangle impactados por diferentes causas vía terrestre, develación de la placa alusiva al proyecto y comida convivio, como cierre del evento.

Fig. 1. Evento inaugural del proyecto “Piloto de restauración ecológica y captura de carbono y emisión de metano en los de manglares de Costa Rica y Benín” con autoridades de Benín y México en la municipalidad de en Ouidah, Benín. África.
Del 16 al 26 de octubre de 2019 se realizó la tercera visita al sitio de restauración en Ouidah (Figura 2). El objetivo principal de esta fue transferir el conocimiento sobre las técnicas de reforestación, colecta de hipocótilos, detección de los sitios críticos por azolve en los canales artificiales. Se impartió la asesoría sobre las técnicas a seguir para el desazolve efectuado en diciembre de 2019 y febrero de 2020 considerando el nivel del agua. Además, se asesoró sobre técnicas de producción de plántulas de mangle (Avicennia germinans y Rhizophora racemosa), estableciendo el tipo de contenedor y preparación del sedimento, definiendo una fórmula para el sitio de restauración. Con la finalidad de dar soporte técnico en ambas actividades, se proporcionó al Biol. Ebénézer Houndinou, responsable técnico de ejecución; una guía técnica para desarrollarlas. Se superviso los trabajos realizados en la colecta e instalación de las cámaras hidropónicas para la producción de plántulas de A. germinans. Cabe resaltar que durante la capacitación de los técnicos de la ONG CORDE y el estudiante de maestría, personas de la comunidad también participaron.
Con respecto a las asesorías en el 2019 del estudiante de maestría, se realizaron reuniones para revisar los protocolos y presentaciones para defensa de avances y tesis, se actualizo la base de datos y se proporcionó una base bibliográfica de artículos científicos desde 2017 a la fecha. Para ello, se creó en una carpeta Google Drive, donde la Universidad Autónoma de Campeche asigno espacio ilimitado.

Fig. 2. Visitas en octubre de 2019 a las comunidades con injerencia al proyecto de restauración de manglares ubicados en la municipalidad de Ouidah, Benín. África.
Se impartió en el 2019 una conferencia sobre el protocolo por aplicar de restauración en Ouidah al Comité Técnico de Humedales, para su aprobación, la cual fue positiva (Figura 3). Asimismo, se realizó una reunión con las comunidades potenciales para los trabajos de excavación, donde se les explico el proyecto y las técnicas a seguir por parte del Biol. Ebénézer Houndjinou y la Dra. Claudia M. Agraz Hernández (Figura 3).

Fig. 3. Conferencia impartida en el 2019 sobre el protocolo por aplicar de restauración en Ouidah al Comité Técnico de Humedales (lado izquierdo) y reunión con las comunidades potenciales para los trabajos de excavación de los canales artificiales (lado derecho); actividades efectuadas por la Dra. Claudia M. Agraz y Biol. Ebénézer Houndjinou.
El monitoreo de los parámetros físicos y químicos de la zona de restauración y bosque de referencia se ha realizado de febrero a diciembre del 2018 antes de realizar las acciones de restauración. Cabe destacar que el mes de junio no presenta datos, pues el nivel del agua no permitió entrar al sitio.
Zona de restauración: el comportamiento del pH registro dos patrones, condiciones ácidas de febrero a julio 2018, con un intervalo de 5.9±0.2 a 6.5±0.1; y ligeramente alcalinas de agosto a diciembre 2018 (de 7.0±0.64 a 7.5±0.5. Figura 4). Asimismo, el grado de oxigenación presenta comportamientos de tipo óxico en febrero (-97.7±35mV) y marzo (-101.2±73mV) (Figura 14), y óxico con tendencia a óxico-hipóxico en el resto de los meses, de -124.2±112mV a -200.9±89mV. Lo anterior, con temperaturas mínimas reportadas en julio de 2018 (27.9±0.2 °C) y máximas en abril de 2018 (29.0±2.73°C. Figura 4). Con respecto a la salinidad la tendencia registrada fue de tipo mesohalina.
Este comportamiento es causado por el menor aporte de agua dulce al sistema durante los meses de febrero a abril registrando mayor demanda de oxígeno, ante los altos tiempos de residencia del agua, la fragmentación del hábitat y aporte continuo de material orgánico; causando un déficit hasta el mes de julio a pesar de darse inicio de las lluvias y el exhibir condiciones óxicas en el agua intersticial. Pues la demanda de oxígeno supera la capacidad de oxidar el nitrógeno reducido (amonio), provocando el almacenamiento del nitrógeno en el sedimento, además de aumentar el amonio en el agua intersticial. Por ello, la concentración del amonio en el agua intersticial de febrero a diciembre, de 2.03 a 13.99 mg/L (Figura 6). Esto sustentado a su vez, con el comportamiento de los parámetros químicos del agua intersticial en los meses que se reportan, con mayor grado de hipoxia, salinidad y acidez hacia la parte interna de la zona de restauración (Figura 4 y 5). Razón por la cual, Sesuvium portulacastrum y Paspalum vaginatum dominan en el área de restauración. Autores como Boto (1984), Lewis (1982), y Mitsch y Gosselink (2000) sugieren que el patrón hidrológico (hidroperíodo) describe las condiciones de óxido/reducción de un sitio específico, estableciéndose con ello cambios en el potencial redox; y, por ende, en los patrones fisiológicos y de tolerancia de las plantas.
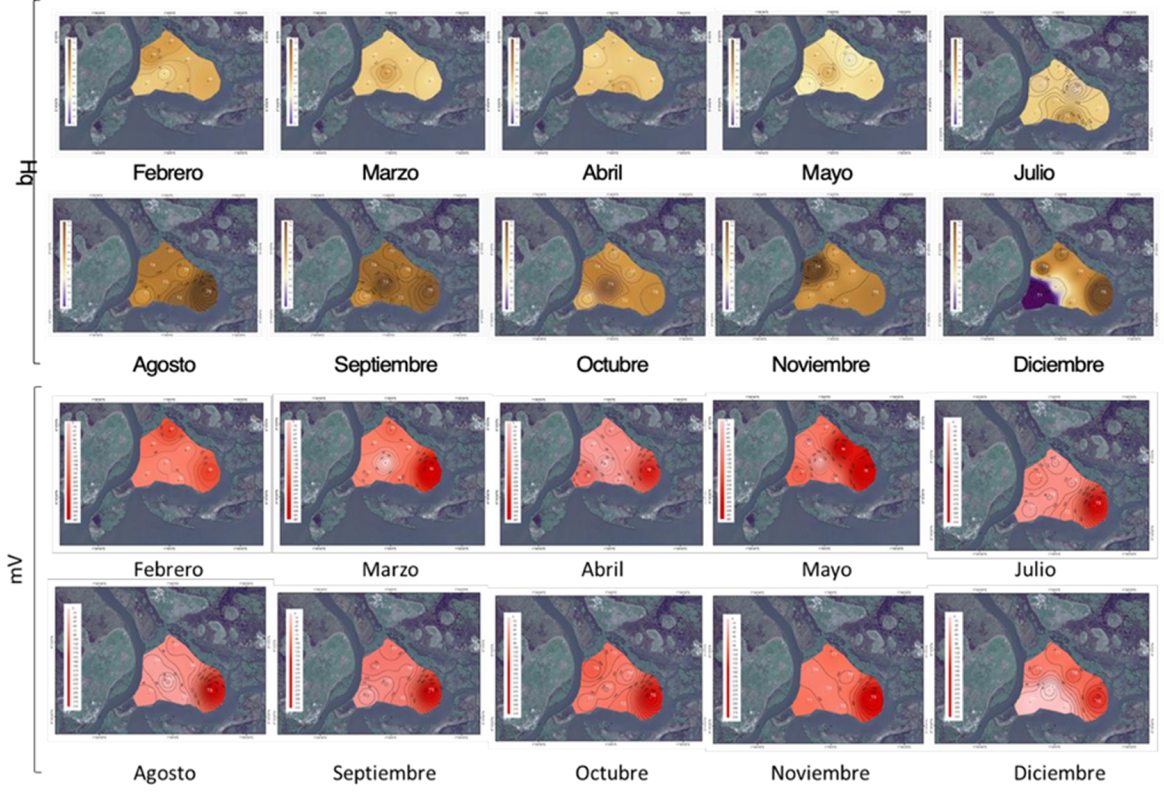
Fig. 4. Comportamiento del potencial redox (mV) y pH en el agua intersticial de febrero a diciembre de 2018 en una zona restaurada de la municipalidad de Ouidah, Benín. África.
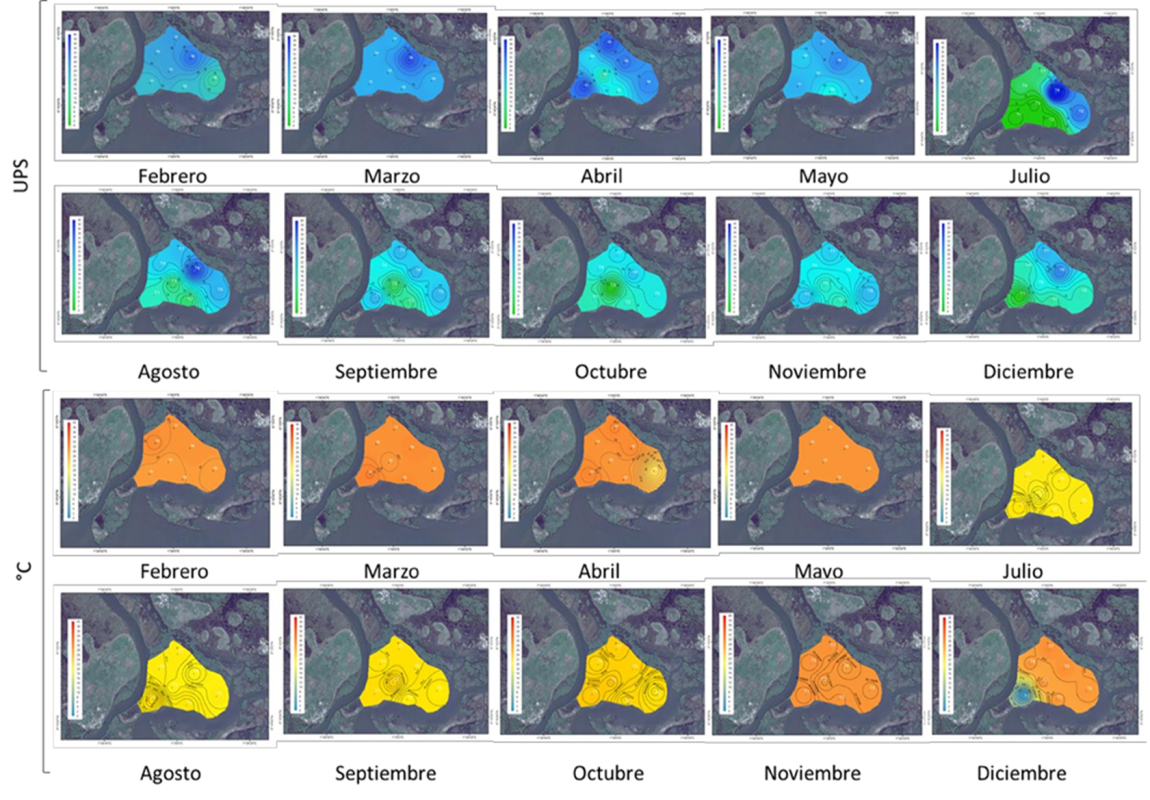
Fig. 5. Comportamiento de la salinidad y temperatura en el agua intersticial de febrero a diciembre de 2018 en una zona restaurada de la municipalidad de Ouidah, Benín.
En general las condiciones tróficas del agua intersticial establecieron antes de realizarse las acciones de restauración entrada de agua residual de origen agrícola y urbanas a lo largo del canal que desemboca al mar acarreados por el desagüe de la laguna Ouedeme, propiciando condiciones hipóxicas tendientes a anóxica en el área de restauración. Esto validado mediante la concentración de compuestos nitrogenados registrados en los meses de monitoreo, así como por el incremento de los aportes de estos y los fosfatos en el mes de mayo en ambos sitios en estudio; he incluso superiores en el área de restauración (Figura 6). Al recibir esta área primero los aportes de agua residual, por acarreo al iniciar las lluvias en el mes de mayo. Rodríguez et al. (2006) mencionan que las actividades socioeconómicas llevadas a cabo en zonas circundantes a los bosques de mangle; pueden generar en mayor o menor grado, estrés e impacto en el desarrollo, evolución y conservación de los manglares.

Fig. 6. Concentración promedio de los nutrientes del agua intersticial durante el periodo de febrero a diciembre de 2018 en el área de restauración ubicada en la municipalidad de Ouidah, Benín. África.
Zona de referencia: El comportamiento general muestran condiciones ácidas de febrero a julio 2018, con intervalos de 5.8±0.2 a 6.8±0.3 y alcalinas de agosto a diciembre (de 7.3±0.8 a 7.7±0.60), similar a lo registrado en la zona de restauración (Figura 7). Sin embargo, las condiciones de óxido reducción definieron condiciones anóxicas de febrero a diciembre del 2018, con intervalos de -328.95 ± 0.49 a -408±1.3 m (Figura 7). Las temperaturas máximas se exhibieron de mayo a agosto de 2018, con un promedio de 29.1±5.0 °C y los mínimos de febrero a abril y de septiembre a diciembre, 27.9±0.7 °C; bajo condiciones en general, oligohalina con tendencia a mesohalinas (16.3 ± 4.3 UPS). Con máximos en marzo (21.3±4.8 UPS) y mínimos julio y diciembre (12.6±5.0 UPS); condiciones que se mantuvieron hasta el mes de junio del 2019 (14.3±3 UPS) (Figura 7).
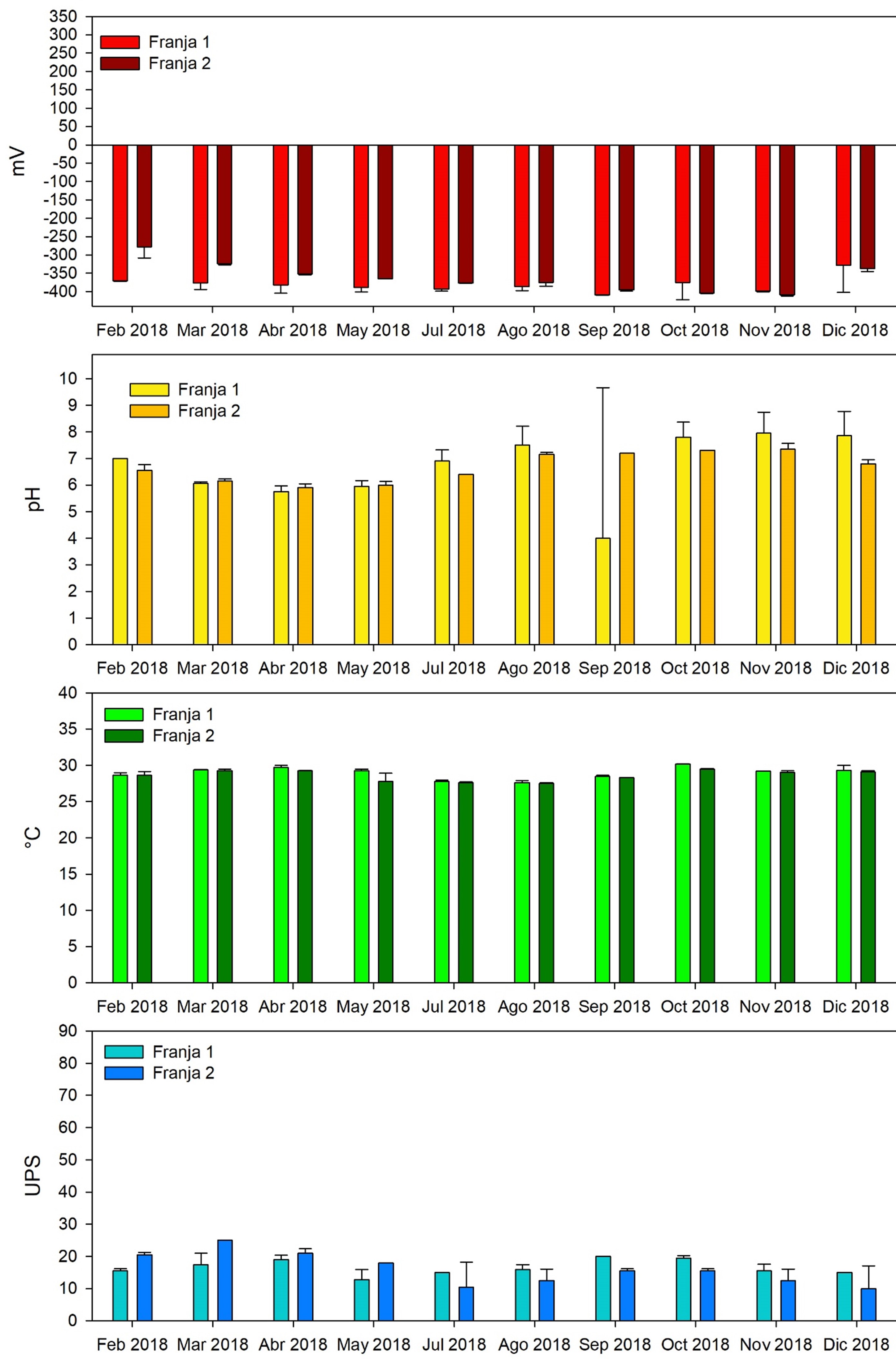
Fig. 7. Concentración y valores de los parámetros físicos y químicos del agua intersticial de febrero a diciembre 2018 en dos bosques de mangle ubicados en la municipalidad de Ouidah, Benín. África. Franja 1: bosque ubicado en la orilla del cauce del río y Franja 2: bosque interno posterior a la franja 1 * Los meses de enero, febrero, mayo y junio de 2019, no fue posible registrarlos, por fallas en el equipo.
El estado trófico del agua intersticial de los bosques de referencia registró mesotrófica, con respecto a la concentración de los nutrientes, comparado con la zona de restauración (Figura 8), principalmente en el amonio, registrándose las mayores concentraciones en octubre (18.8±0.1 mgL-1) y noviembre (35.8±9.0 mgL-1). Esto debido al aporte de agua residual urbana y actividades productivas que se llevan a cabo desde la zona oeste y este. Por esta razón, las condiciones de óxido reducción fueron hipóxicas con tendencia anóxicas (Figura 8). Mohamed et al. (1998) registraron similares concentraciones de fosfatos (0.1 a 2.7 mgL-1) con las concentraciones más altas de los nutrientes en los boques internos; atribuidos al aporte de los nutrientes de origen antrópico en la bahía de Chwaka Zanzíbar en África. Por su parte, Boto y Wellington (1984) mencionan que la variación en los valores del potencial redox en el agua intersticial, es debido a que estos dependen de la frecuencia y duración de inundación de la marea, contenido de materia orgánica y disponibilidad de aceptores de electrones; tales como NO3-, Fe+3, Mn+4 y SO4-2. .
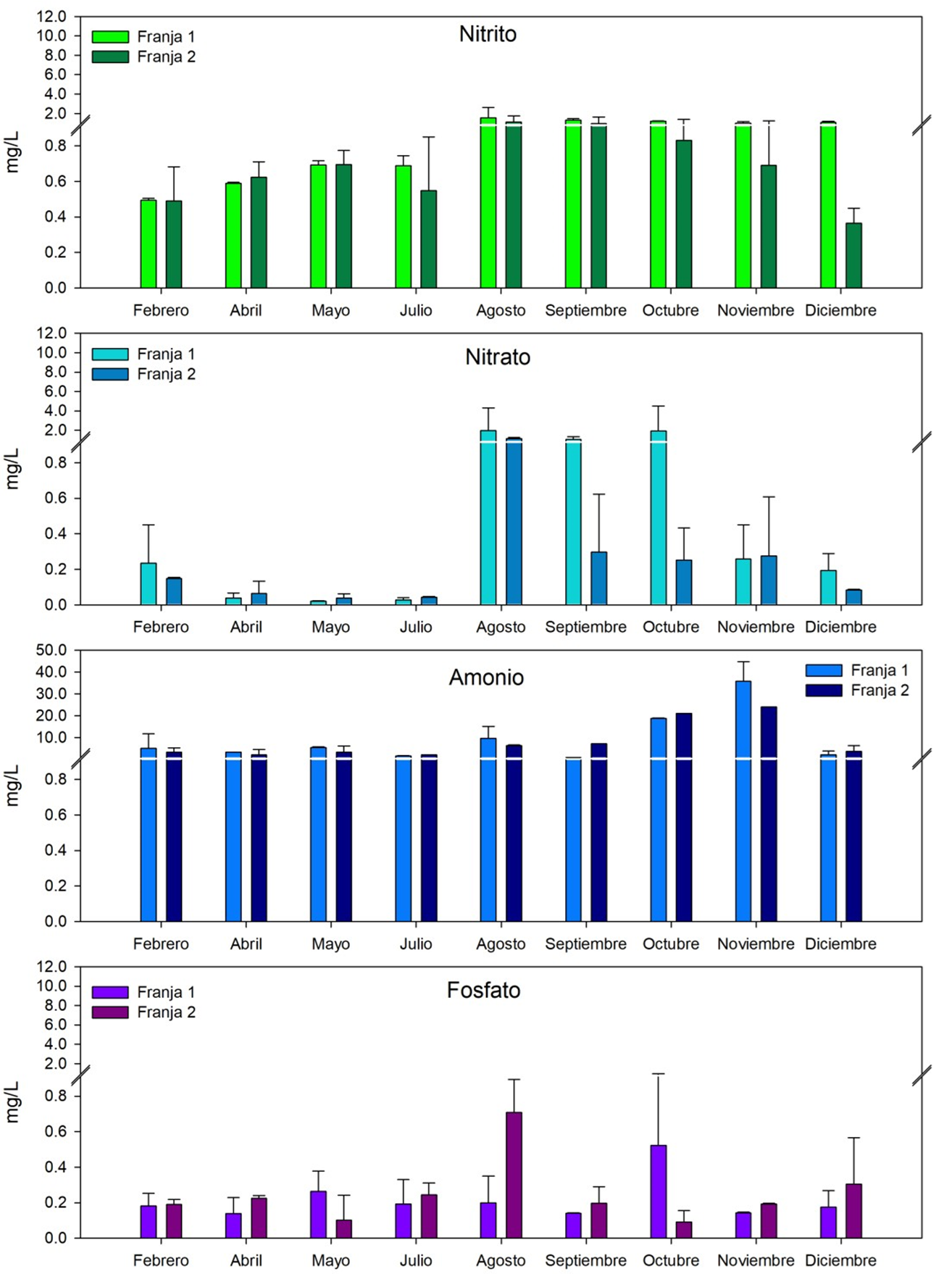
Fig. 8. Concentración de nutrientes en el agua intersticial de febrero a diciembre del 2018 en los bosques con dominancia de Rhizophora racemosa (Franja 1) y Avicennia germinans (Franja 2), ubicados en la municipalidad de Ouidah, Benín. África.

Fig. 9. Concentración de los sulfatos en el agua intersticial de febrero a diciembre de 2018 en los bosques con dominancia de Rhizophora racemosa (Franja 1) y Avicennia germinans (Franja 2), ubicados en la municipalidad de Ouidah, Benín. África.
Se continuo con el monitoreo de los parámetros fisicoquímicos en la zona de restauración y bosque de referencia después de las acciones de rehabilitación hidrológica, efectuadas en el mes de marzo del 2019. Cabe destacar que en el mes de junio no presenta datos, pues el nivel del agua no permitió entrar al sitio. En los meses de enero y febrero del 2019 y el periodo de mayo a octubre del 2019 se realizaron registros de pH, temperatura y potencial redox, pues el equipo presento fallas en los sensores.
Zona de restauración: : el pH incremento ligeramente a alcalino en el año 2019, de 7.3±0.43, posterior a la rehabilitación. Esta tendencia establece mayor entrada del agua de mar al sitio de restauración a través de la excavación de los canales artificiales, sin embargo, no se definió diferencias significativas entre el año 2018 y 2019 (F1,152 = 3.76, p≤0.025; Figura 10). El grado de oxigenación del agua intersticial fue similar antes y después de las acciones de rehabilitación, con -135.9±96.0 mV en el 2018 y de -153.3±93.2 mV para el 2019, respectivamente (F1,151= 142.39, p≤0.0001; Figura 11), a pesar de la gran cantidad de materia orgánica suspendida durante la excavación de los canales artificiales.
Similar comportamiento se registró para la temperatura entre los años 2018 y 2019 (F1,152 = 1.52, p≤0.221; Figura 10), así como entre el 2020 y 2021 fue igual, definiéndose un valor promedio para los cuatro años de 28.9±1.1 °C (Figura 10). Mismo comportamiento fue detectado en general las condiciones óxicas del 2018 al 2021, de -155.5±30.7 mV (Figura 11 y 12).
Caso contrario con el comportamiento de la salinidad que exhibió diminución en la concentración desde el 2018 al 2021, con 21.3±3.7 a 11.1±3.4 UPS (Figura 10 y 11) Con respecto a la salinidad la tendencia registrada fue de tipo mesohalina durante el 2018, periodo antes de las acciones de rehabilitación hidrológica, de 21.3±7.5, con disminución en la concentración de la salinidad durante el 2019, con 13.4±2.7 UPS estableciéndose diferencias significativas (F1,207 = 26.95, p≤0.001; Figura 11). Posterior a este periodo se establece el incremento de 3 UPS en el 2020, causado por el déficit de 13.2% de la precipitación. A pesar de esto, la salinidad se registró en el 2021 por debajo de la concentración establecida antes de la rehabilitación hidrológica (10.2 ± 0.3 UPS; Figura 10).
Con respecto al pH, este se incrementó desde condiciones ácidas (6.7±1.2) en el 2018 hasta alcalinas (8.1±0.8) en el 2021. Los cambios registrados en la alcalinidad, en conjunto de la recuperación del comportamiento del hidroperíodo, ha generado la disminución en un 80% Paspalum vaginatum en el área de restauración, incrementando la regeneración natural de R. racemosa en un 15% y el resto del área con Sesuvium portulacastrum.

Fig. 10. Comportamiento de la salinidad y temperatura en el agua intersticial enero a diciembre de 2019 en un área restaurada ubicada en la municipalidad de Ouidah, Benín. *Sin datos en enero, febrero y de mayo a octubre por fallas en el equipo.
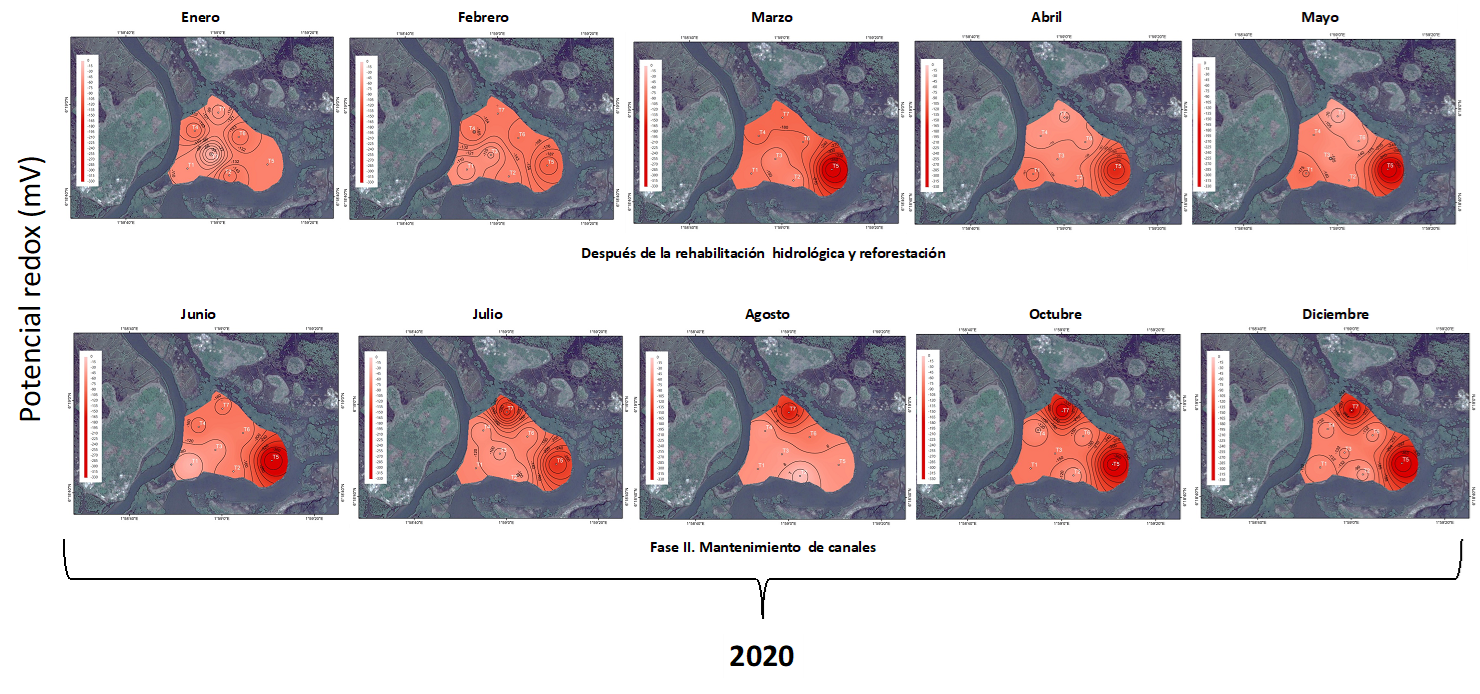
Fig. 11. Comportamiento del potencial redox en el agua intersticial de una zona restaurada del Municipio de Ouidah, Benín *Sin datos en enero, febrero y de mayo a octubre por fallas en el equipo.
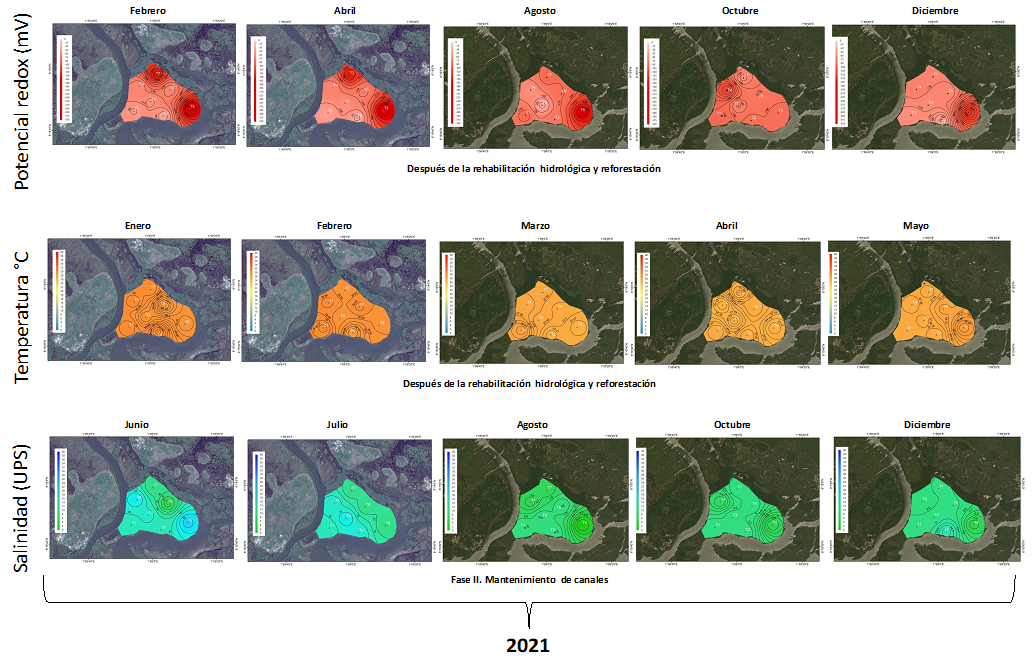
Fig. 12. Comportamiento del potencial redox, pH, temperatura y salinidad del agua intersticial de enero a diciembre de 2021 en un área restaurada ubicada en la municipalidad de Ouidah, Benín. África.
Bosque de referencia: el comportamiento de los parámetros físicos y químicos del agua intersticial del 2019 al 2021, fue similar a los registros en la zona de restauración. Para el 2019 las condiciones ambientales se mantuvieron similar al 2018, sin embargo, para los años 2020 y 2021, el pH exhibió alta alcalinas , manteniendo condiciones hipóxicas a anóxicas (-245.0±90 mV a -417.8±59.1.7 mV)(Figura 13). En el caso de la salinidad disminuyo de 16.6±2.3 a 8.9±5.4 UPS, a pesar del 38%déficit de la lluvia (Figura 14).
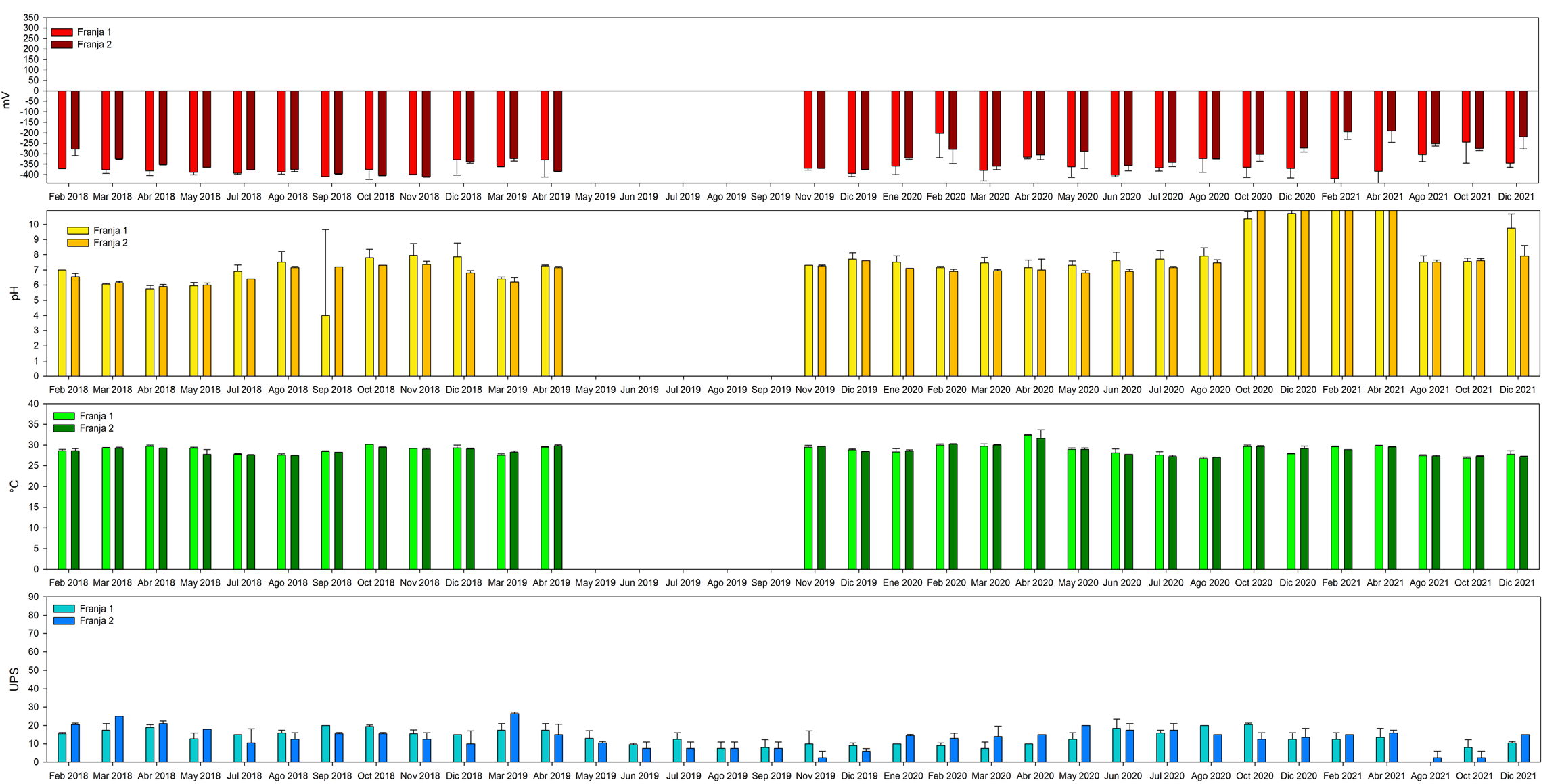
Fig. 13. Comportamiento de los parámetros físicos y químicos durante y posterior a las acciones de restauración en unos bosques de mangle desplazado por Paspalum vaginatum ubicado en la municipalidad de Ouidah, Benín. África.
La precipitación mensual de enero de 2018 a abril de 2022 de la estación meteorológica de Cotonou, de la estación meteorológica: 653440(DBBB) (https://www.tutiempo.net/clima/12-2018/ws-653440.html), más cercana al área de restauración (Figura 12). Esto con la finalidad de elaborar el climograma para identificar el tipo de clima, como este influye en las diferentes físicos y químicas del agua intersticial en el tiempo y su relación con la producción de hojarasca. Al comparar los periodos de enero a diciembre del 2018, con respectivo al año 2019, se establece 7.2% más de lluvia en el año 2019, sin embargo, se registra el desplazamiento del comportamiento de las lluvias. Al determinarse la época de mayor precipitación de mayo a octubre en el 2018, caso contrario en el 2019 donde las máximas precipitaciones se registraron en dos periodos, de enero a junio y setiembre y octubre, con registros de sequía en el mes de agosto del 2019 (Figura 14). Similar comportamiento fue registrado en los Humedales de Térraba Sierpe en Costa Rica (consultar este documento). Asimismo, se registró la disminución del 31% la precipitación comparando del año 2019 al 2020, con aumento en la temperatura ambiental de 2.4 oC en el primer periodo de años, en marzo de 2020 respecto al mismo mes del año 2019. Del 2020 al 2021 la precipitación disminuyo el 3.4%, modificándose la estacionalidad (Figura 14). El déficit y cambios de la estacionalidad en la precipitación es atribuida a los efectos del cambio climático que están incidiendo en los ambientes costeros principalmente en los patrones de temperatura y precipitación. Similares resultados fueron registrados por Agraz Hernández et al. (2017) y Chan Keb et al. (2018) en los manglares del Estado de Campeche, al establecer cambios atípicos interanuales en la precipitación y entre las épocas del año, durante el periodo del 2006 al 2009, como reflejo de la variabilidad climática a nivel global.
Diversos autores indica que los efectos del cambio climático propician modificaciones en la temperatura del aire y la precipitación, aportes de agua dulce, la circulación oceánica y atmosférica, y la tasa de elevación del nivel del mar; así́ como la estacionalidad y distribución de los huracanes y las tormentas tropicales (Blasco et al., 1996; Scavia et al., 2002).
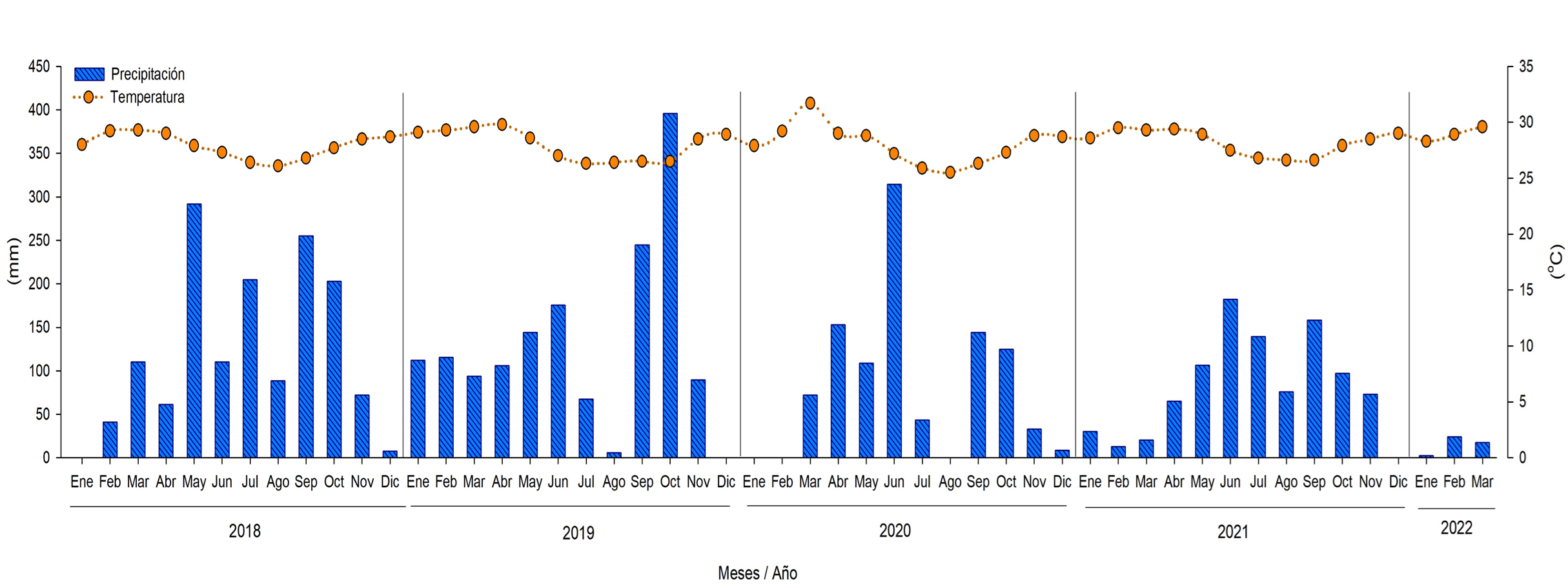
Fig. 14. Precipitación total de enero de 2018 a abril de 2022 en el Cotonou, Benín, África. Datos reportados por la estación meteorológica: 653440 (DBBB). Latitud: 6.35 | Longitud: 2.38 | Altitud: 5.
Para establecer la producción de hojarasca y fenología en los bosques de referencia, se instalaron 6 canastas de defoliación en cada bosque (Figura 15). El monitoreo, procesado y conservación de las muestras se efectúan mensualmente, por especie. Este trabajo está efectuando cada mes por CORDE, con las metodologías aprendidas en el febrero del 2018, mediante la capacitación de los investigadores de México. En el mes de junio del 2018, durante la estancia de los investigadores de México, se efectuó la capacitación del procesado de la información, para ser interpretado en el tiempo.

Fig. 15. Instalación de las canastas de defoliación en el bosque de Avicennia germinans (referencia).
La determinación de producción de hojarasca, indica de manera indirecta el aporte de la biomasa vegetal que se integra a la fuente de nutrientes para diversos organismos que habitan en los manglares y ecosistemas adyacentes. Pese a que los manglares son plantas que defolian sus hojas durante todo el año, tienden a incrementar la caída de hojarasca en alguna época del año, en específico lluvias o durante la sequía prolongada (Flores-Verdugo, 1985; Day et al., 1996; López- Portillo y Ezcurra, 2002 y Agraz Hernández et al., 2011). Asimismo, las diferentes especies y tipos fisonómicos producen diferente biomasa vegetal, tal como fue observado en este estudio.
La mayor producción de la hojarasca en los bosques de referencia Rhizophora racemosa aportó más del 30%, comparado con Avicennia germinans (Figura 16). La menor productividad en A. germinans puede ser atribuido al mayor grado de estrés que recibe el humedal por asentamientos humanos, deforestación, aportes de agua residual, fragmentación del humedal (cambiando el patrón hídrico) y por la dominancia en el flujo de agua laminar; favoreciendo el incremento en la salinidad, temperatura del agua y condiciones hipóxicas a completamente anóxicas en el agua intersticial. Así como, el cambio hidrológico que se ha venido realizando a través del cambio de la microtopografía, derivado de la extracción de sal. Lichtenthaler (1996) indica que los gradientes de salinidad son factores que regulan los procesos fisiológicos en los manglares y establecen que en las zonas donde se encuentran manglares con alta concentración de salinidad, presentan estrés fisiológico y, por ende, reducción en la asimilación del dióxido de carbono, en la conductancia estomática y las tasas de crecimiento.
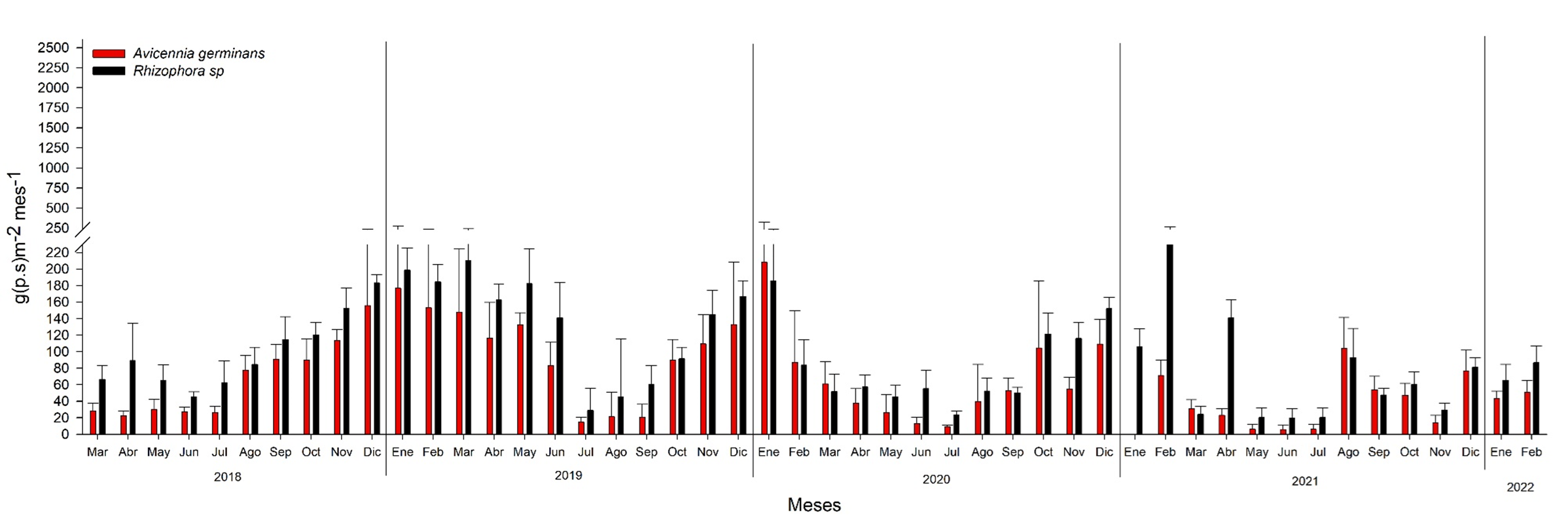
Fig. 16. Producción de hojarasca mensual en dos bosques de mangle monoespecíficos de Rhizophora racemosa (Franja 1) y Avicennia germinans (Franja 2) ubicado en la municipalidad de Ouidah, Benín. África.
En general la época de floración fue de agosto a diciembre para R. racemosa y de julio a noviembre para A. germinans (Figura 17 y 18). En el caso de la producción de propágulos se definió de julio a diciembre para ambas especies; información relevante para la producción de plántulas de A. germinans y R. racemosa, así como la reforestación in situ con hipocótilos de R. racemosa, que se llevó a cabo entre octubre y noviembre del 2019, en la zona de restauración (Figura 17 y 18). Esta estacionalidad es inducida por los parámetros medio ambientales y a las características particulares de cada especie, tales como; el periodo de producción de frutos y flores, la duración de vida de las hojas y el tiempo de desarrollo de la flor hasta ser un propágulo.
Para el año 2019 se incrementó la producción de hojarasca, con 1618.2 g.m2año-1 en el bosque de R. racemosa y 1200.0 g.m2año-1 para A. germinans, debido al incremento en la precipitación de 2018 al 2019. Sin embargo, al comparar la producción de 2019 al 2020 de ambos bosques de enero a mayo disminuyo en un 73.5 a 82.9%, como consecuencia de la menor precipitación (-31%) durante este periodo; afectando la reproducción fenológica, con menor producción de flores y propágulos en A. germinans y R. racemosa. Para el 2021, debido al mayor estrés que se generó ante la disminución del 3.4% de la precipitación de este año, con respecto al 2020, la producción de hojarasca disminuyo 12.5% más, se mantuvo baja producción de flores y propágulos, e incluso en A. germinans se produjo 79% y 69.3%, respectivamente, comparado del 2019, con respecto al 2020 y 2021.
En general los bosques de R. racemosa y A. germinans están definidos con base a la producción de las hojas; como “jóven”, pues la producción del tejido fotosintético corresponde en el periodo de los 4 años monitoreo, más del 60% comparado con el resto de los componentes de la hojarasca (Figura 17 y 18). Este aporte coincide con el patrón de productividad observado por Lacerda et al. (2001), Aké-Castillo et al. (2006), Agraz Hernández et al. (2011).
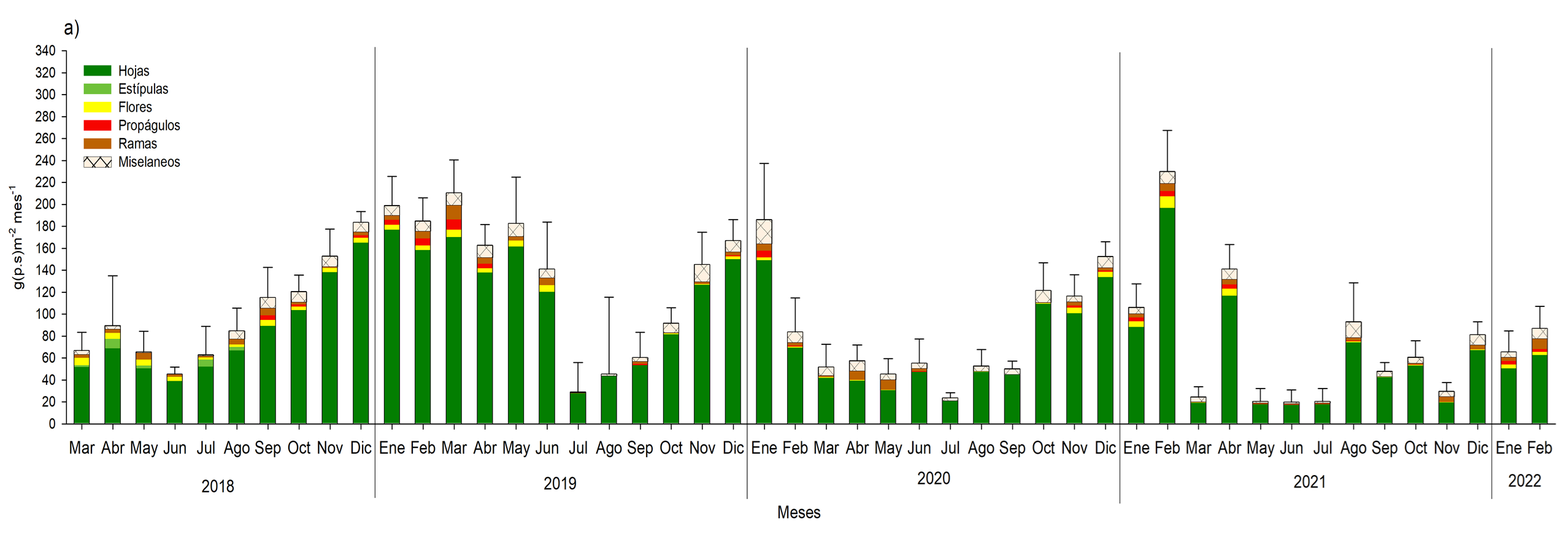
Fig. 17. Aporte de hojarasca de marzo de 2018 a febrero de 2022 en un bosque monoespecíficos de Rhizophora racemosa, ubicado en la municipalidad de Ouidah, Benín. África.
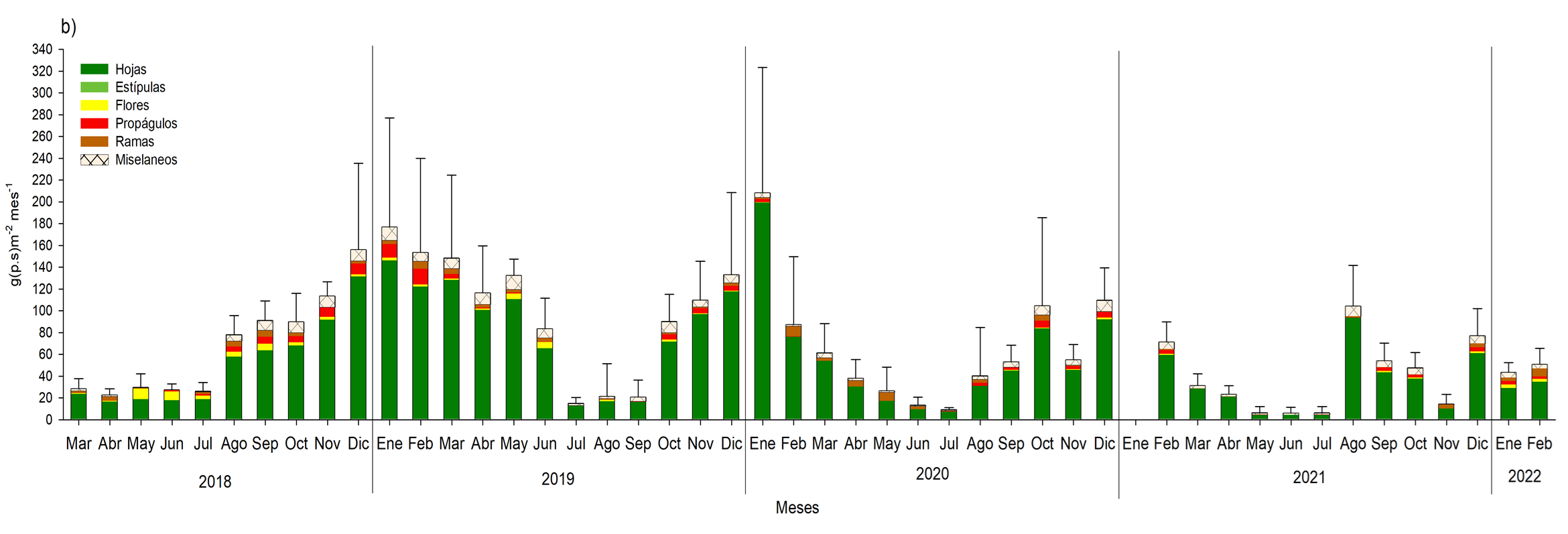
Fig. 18. Aporte de hojarasca de marzo de 2018 a febrero de 2022 en un bosque monoespecíficos de Avicennia germinans, ubicado en la municipalidad de Ouidah, Benín. África.